

What I listed while I wrote this article.
It is not every day that a paper is published that gets me excited enough to break my schedule, it did happen a few times in the last 2 years, but it is becoming rarer and rarer. As I previously stated, the low-hanging fruits are all picked and what is left now are only very complex puzzles, waiting to be solved.
While proposing a central “hypothesis”, this hypothesis is made up of many pillars. The Spike interaction with dozens of receptors leads to a caleidoscope of pathways being activated, the Spike interaction with biofilms and endotoxins, the Kynurenine Pathway and consequential hijacking by the virus/Spike, and many other aspects of this led to what I once named “Reverse AIDS”, now called Paradoxically Acquired Immune Dysfunction…or PAID… truthful and hilarious at the same time.
But among the pillars of this hypothesis lies molecular mimicry. In simplistic terms, how very small pieces of the virus mimic pieces of other pathogens, other things, which were then able to induce some sort of reaction (disease). For a short while, I considered naming this Substack “Plagued by Design” precisely because of this.


The second one bears a lot more weight to our discussion here. The Spike Protein Receptor-Binding Domain is riddled with molecular mimics of other pathogens, a specific section of the Spike mimics a transporter of tryptophan potentially leading to autoimmunity against it, hindering this important function, thus leading to immune dysfunction. Being the center of attention for a good few years, it was found the Spike is in fact, a great mimic.
These small pieces of the virus that can cause all of these effects are often called “viral fragments”, another pillar of my central hypothesis. Persistence of small pieces of the virus inducing inflammation long-term. Leading us to this exciting paper.
Highlights of this awesome paper.
Small pieces (fragments) of SARS-CoV-2 resemble small peptides that we produce to fight infection
These small fragments named xenoAMPs can interact with our own peptides and create even more inflammation
These fragments are created when own body breaks down the virus
Viral afterlife: SARS-CoV-2 as a reservoir of immunomimetic peptides that reassemble into proinflammatory supramolecular complexes
While we now have a deep understanding of coronaviruses, but especially how SARS-CoV-2 induces disease, there are still many unanswered questions, many pieces of the puzzle that have yet to be elucidated. Among these is why SARS-CoV-2, unlike other coronaviruses, such as the common cold, can induce severe disease, cytokine storms, organ damage in organs without any sign of the virus, all the different clotting, the unique skin presentation, high levels of autoantibodies.
The authors raise the same point I did multiple times, what happens when the virus is broken down is smaller fragments, given the numerous small fragments that resemble “other things” and can initiate a myriad of reactions. So could the breakdown of the virus by the immune system create small fragments that mimic potent peptides ?
Peptides → small quantity of amino acids together
Proteins → large quantity of amino acids together
By using Machine Learning the authors used their model to analyze the entire virus protein sequence and found, yes, certain small fragments of the virus mimic antimicrobial peptides (AMPs), small peptides that can kill bacteria, viruses, and fungi, the ones found in SARS-CoV-2 are called XenoAMPs.
They chose to compare the possible xenoAMPs to the human cathelicidin LL-37, one of the most potent AMPs we have, because it has a high cationic (positive) charge, and it can organize and form functional structures with dsRNA (which is negatively charged), something released from damaged cells when we get infected.
Both Neutrophil Elastase and Matrix Metalloproteinase-9 (they break down proteins) are capable of generating these xenoAMP. Compared to the common cold coronavirus (HCoV-OC43) which is very mild, SARS-CoV-2 has higher scores
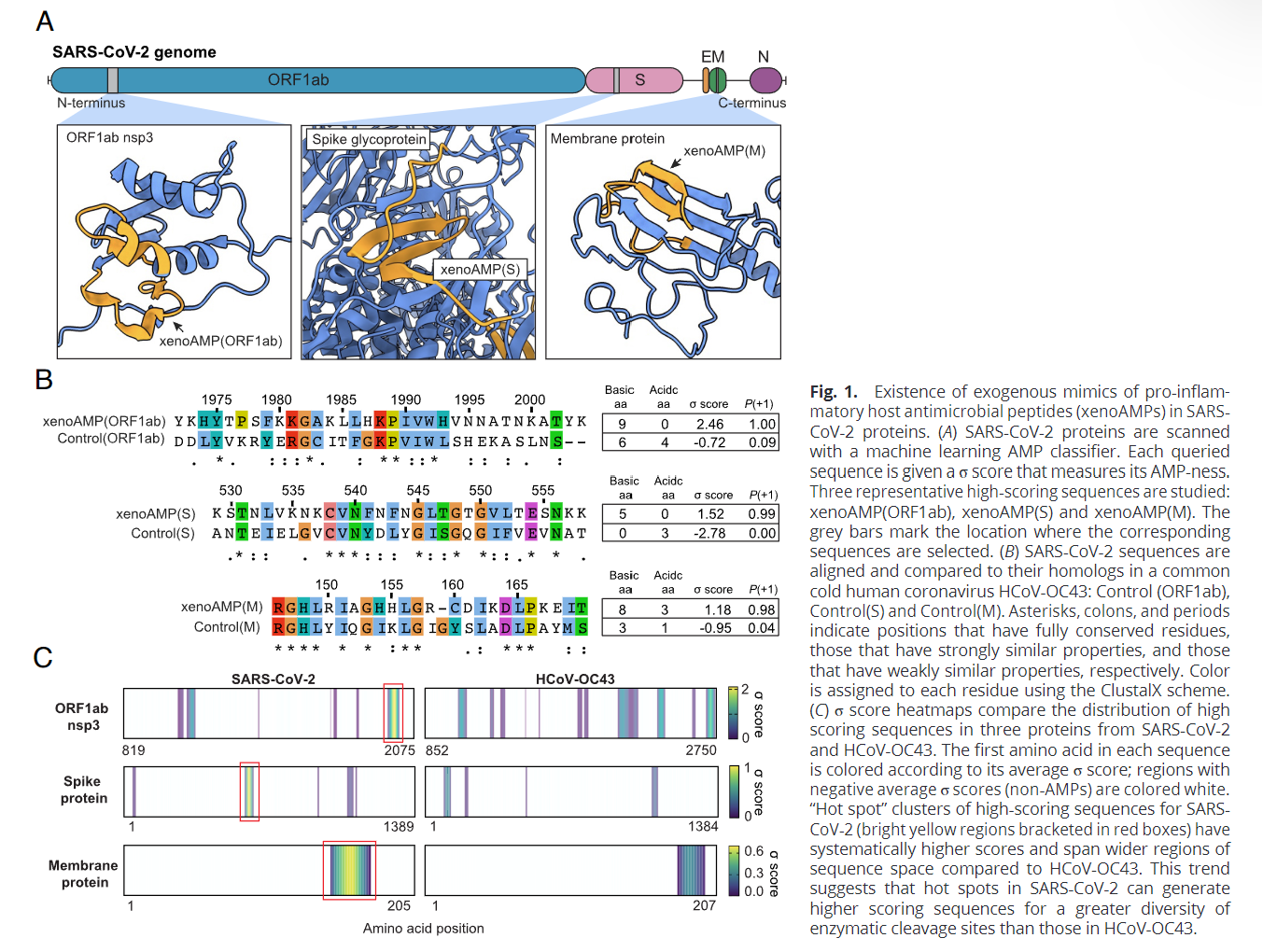
Besides how the protein behaves with water (hydrophobicity), the charge is the other most important aspect of it. XenoAMPs having higher cationic (positive) charge is one of the reasons it forms proinflammatory structures with dsRNA which are anionic (negatively charged) because it is how we “fight infections”.
In most cases, free AMPs (imagine it floating around your body) aren’t expected to be found in high concentrations in the body. By checking 29 severe COVID patient samples, they found LL-37 fragments in 20 of 29 samples, but on the other hand, 28 out of 29 samples had SARS-CoV-2 fragments, with some of these being XenoAMPs.
To valid their machine learning and observations so far, the authors used fairly intricated testing, and observed that the xenoAMPs organize (glue together) with dsRNA in a similar manner as the LL-37, a human AMP, even though xenoAMPs are not as well-structured as AMPs (remember, the shape and structure of proteins are very important).
When LL-37 “glues” itself with dsDNA it forms a structure that can be recognized by TLR3 and TLR9, thus potentializing the immune activation by a large margin. To test if the same happens with xenoAMPs, they used poly(I:C) to mimic viral dsRNA produced during viral replication, and found that indeed xenoAMPs form similar complexes that can induce a strong immune response. Remarkably enough, they also tested and found that SARS-CoV-2 xenoAMPs can form functional structures (cocrystallize into the same columnar lattice) with LL-37 suggesting that xeno AMPs and human AMPs can work synergistically together.
Comparing SARS-CoV-2 xenoAMPs with their weaker homolog in the common coronavirus cold they found that xenoAMPs+synthetic dsRNA (polyI:C) release more inflammatory proteins than their respective common cold cousin. The lungs of SARS-CoV-2-infected people have substantial amounts of RNA, which could, in theory, amplify the inflammation in vivo.
Further testing in different cell lines showed that xenoAMP+poly(I:C) complexes had a distinct higher expression of inflammatory genes CXCL1, IL8, CXCL10, IL1b, IL6, GCSF (granulocyte colony-stimulating factor), IFNB1, and others.
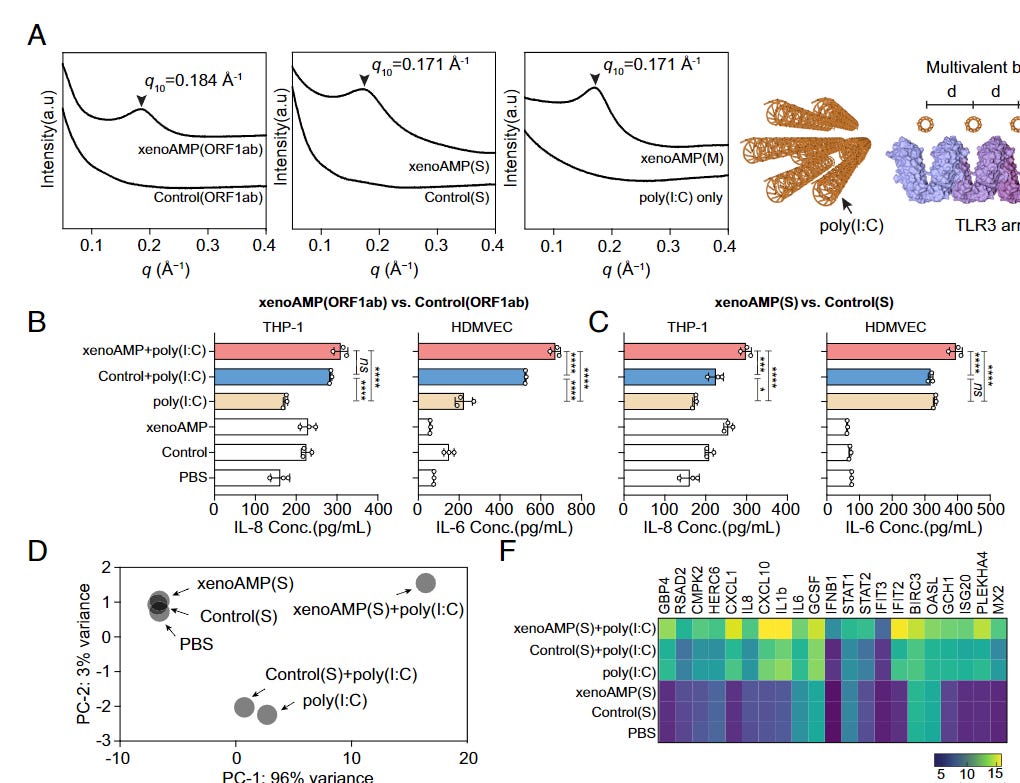
Testing in many different cell lines, mostly to do with the endothelium and skin demonstrated the same, the combination of xeno+poly induced the production of more inflammation that was in line with the same level of inflammation you can see in SARS-CoV-2 infection, including another potential mechanism for coagulation amplification.
As their last step, they went to measure the capacity of xenoAMPs+poly(I:C) complexes to induce immune activation in vivo using a mouse model.
Compared to poly(I:C)- only treatment, xenoAMP(ORF1ab)- poly(I:C) treatment strongly increases plasma IL- 6 and CXCL1 levels by 1.6× and 2.2× respectively. Immune activation by xenoAMPs- poly(I:C) complexes is evaluated specifically in the lung, which is strongly affected in COVID-19 patients (41). CXCL1 and IL- 6 levels due to xenoAMP(ORF1ab)- poly(I:C) increase by 1.2× compared to poly(I:C)- only stimulation. (Fig. 4B) CXCL1 is a chemoattractant for neutrophils, the infiltration of which in the lungs can drive ARDS in COVID-19. IL-6 is a proinflammatory cytokine, and high plasma levels of IL-6 serve as a strong predictor for nonsurvival in COVID-19 patients (7). Importantly, xenoAMP(ORF1ab)- poly(I:C) complex significantly increases the cell count for white blood cells (WBCs), which may be related to the enrichment of neutrophils, lymphocytes, and monocytes (Fig. 4C). These results also suggest that inflammatory responses are xenoAMPs- dependent and heterogeneous, even for genetically identical mice, and that future in vivo comparisons using xenoAMPs from the many new variants and the many nonpandemic coronaviruses may be fruitful.
The findings in the mouse model reinforce all their other observations and testing. xenoAMPs, especially when forming complexes with the proteins inside us not only can induce an inflammatory response, but these complexes will also enhance this inflammatory response, causing a substantial increase in damage.
The importance and significance and how this fits in the bigger picture

A quote, directly from the article above.
No person on the planet (except perhaps Leidos…) has any idea about the proteins and peptides being produced by the mRNA, nobody has any idea about the structure if there are added glycans, amyloid fibrils, or anything else, but above all else, nobody knows or is even testing the possibility and physiological function of the cryptic peptides (non-canonical peptides), and these sequences will have entirely different functions from the protein they originated from (especially if there are added molecules to the protein produced by mRNA).
You do not need a large amount of these peptides to have profound physiological effects. At this point, as a friend wrote to me, we are purely into experimental science without regard to anything else. There is no argument if this will backfire, the only honest argument is far this will backfire.
While this holds true to this day in regards to the mRNA vaccines, for the longest time I have written again, and again, that the virus itself had cryptides, otherwise referred to as “non-canonical peptides”, “small proteins” that are functional, have an effect on our bodies and we are completely unaware.
These peptides will often show themselves either when the protein changes “shape”, but mainly when it is broken down by the body. As I wrote recently, “misfolded” proteins such as amyloid have a complex purpose, they even work as somewhat of a last-ditch effort against infections, acting as a backup antimicrobial peptide.
Here is the catch, AMPs are protease-resistant. They are much harder to break down than other proteins, because of their nature and how they interact with such proteases. This effectively means some of these xenoAMPs may be able to mimic the behavior of “misfolded” proteins and further resist degradation, thus leading to persistent inflammation in the body.
This paper also explains why there is such a massive discrepancy between the vast majority of mild cases and severe infection, since the breakdown of proteins is vastly unique, it will vary even between cells in the same person.
Given how these xenoAMPs can interact with our AMPs (LL-37) and dsDNA, it is highly significant to keep complex interactions in mind. LL-37 has a role in certain autoimmune diseases, especially skin-related ones, and Lupus too (I guess this time, it was Lupus).

Many different pathogens play in neurodegeneration, LL-37 is also implicated in that regard. “Given AD’s association with infection and that overloading AMP may exacerbate AD, this study suggests that LL-37, which is up-regulated upon infection, may be a driving force behind AD by acting as an endogenous agonist of CLIC1.” LL-37 is a binding partner for Amyloid Beta and can impact Alzheimer’s initiation and progression to both ends (stop, or start it). It has a direct role in neurological inflammation.
LL-37 also interacts and modulates, deeply how mast cells act and function. It can interact and prime platelets to induce tissue inflammation and thrombosis. Overexpression of LL-37 is a dual-edged sword when it comes to cancer development. It can promote the development and acceleration of ovarian, lung, and breast cancer and suppress colon and gastric ones.
There is a lot more complexity underlying the findings here and the potential they have, but as a closing remark, I will leave just an image. Nothing else. After all. All roads lead to Rome.

Your support is appreciated and helps my work !

There are more hidden stuff inside this virus.
Just some comments that don't really contribute anything vital here.
Your points about the need for the availability AI are relevant, given that the complexity here is so far beyond mere human ability to keep up with what needs to be learned. Also, standard healthcare has long been over its head with regards to already existing complicated health problems, given the primary goal of waiting for one new drug to address a constellation of symptoms. I have no faith in the ability of standard healthcare practitioners to even diagnose people with the types of issues that you describe, never mind offering effective treatments. That isn't going to change for a long time. The patient's symptoms will be denied along with the MDs' ignorance, resulting in non-treatment at best, and abusive gaslighting at worst.
This also makes me think of the growing “there was no novel virus” crowd. The phenomenon of iatrogenic harm/death does not prove the non-existence of a pathogen. “Where are the vast death numbers then?” is the rhetorical question, and the response is: “given what is described, vast numbers of death are not what is expected. It's an insidious, slowly evolving plague with such a vast array of characteristics that makes it impossible to pin down for those who are using a narrowly targeted pin.”
As an aside, it reminds me of toxic brain damage. It's too defuse and small to be picked up by scans, and can only be discovered via neuropsychological evaluation. Which reminds me of one of the issues that some people are warning about the prolonged use of Ivermectin. The type of brain damage that they describe is highly unlikely to be found, but it's there and it can greatly interfere with people's ability to function as they used to. There's will be “no evidence” of illness and gaslighting will ensue. The point here is not that Ivermectin doesn't have its place; it's the cavalier use of it that we're seeing that's the potential problem.