
The intricacies of T-Cell exhaustion
And SARS-CoV-2, and myocarditis

Announcement - There is an important message from me, pinned in the comment section of this article, please read it.
Well, before delving into the central theme of this article, here are a few clarifications and a summary. This particular paper has been among my sea of tabs (I have roughly 700 tabs among 3 devices, an old laptop, tablet, and my mobile), but covering this particular paper before “building” the overall theme and dynamics in regards to SARS-CoV-2 would make it somewhat meaningless. Another piece lost “in the middle” like a couple of dozen of other articles I published. Although significant, they are lost, like tears in rain.

As a matter of fact and for the sake of clarification, the topic of T-Cell exhaustion remains contentious even among immunologists, PhDs, and other members of the scientific community, there is currently no consensus, which in my opinion is beneficial for the exploration of many possibilities. So what I will present here is but one perspective, of many others. One pertinent argument and also a point of debate is how to distinguish “dysfunction” and exhaustion, at what point the first becomes the second.
Summary of previous articles
Over the past several weeks, I have extensively covered various components of a complex cascade of events often triggered by SARS-CoV-2 (virus or mRNA). These events encompass crucial interactions leading to sepsis, latent infections, metabolic traps (such as the Kynurenine Pathway and IDO), and mitochondrial dysfunction. All of these topics, explored in previous weeks, have the potential to contribute to T-Cell Exhaustion, surpassing mere immune dysfunction. Indeed, if we were to thoroughly connect the dots, it is likely that at least 35% of the content I have covered contributes to the phenomenon of exhaustion. This topic is remarkably extensive in its own right.
The many metabolic and immunological traps written about recently all play different roles in what we will discuss, in fact, if one was to extensively connect the dot, one could probably link at least 35% of what I have written into contributing towards the exhaustion. It is an absurdly extensive topic on its own.
T-Cell Exhaustion in a very simplistic term is the following :
In certain cases, prolonged immune responses or particularly strong infections/reactions to stimuli can result in overworked immune cells, particularly T-Cells. The continuous stimulation and effort exerted on these cells can lead to a state of overwhelm, formally known as "exhaustion." In this exhausted state, T-Cells progressively lose their effectiveness and ability to combat pathogens and maintain bodily clearance of these pathogens.
All credits go to the author, Kevin Roe for this brilliant paper.
Concurrent infections of cells by two pathogens can enable a reactivation of the first pathogen and the second pathogen's accelerated T-cell exhaustion
When multiple intracellular pathogens, such as viruses, bacteria, fungi and protozoan parasites, infect the same host cell, they can help each other. A pathogen can substantially help another pathogen by disabling cellular immune defenses, using non-coding ribonucleic acids and/or pathogen proteins that target interferon-stimulated genes and other genes that express immune defense proteins. This can enable reactivation of a latent first pathogen and accelerate T-cell exhaustion and/or T-cell suppression regarding a second pathogen. In a worst-case scenario, accelerated T-cell exhaustion and/or T-cell suppression regarding the second pathogen can impair T-cell functionality and allow a first-time, immunologically novel second pathogen infection to escape all adaptive immune system defenses, including antibodies. The interactions of herpesviruses with concurrent intracellular pathogens in epithelial cells and B-cells, the interactions of the human immunodeficiency virus with Mycobacterium tuberculosis in macrophages and the interactions of Toxoplasma gondii with other pathogens in almost any type of animal cell are considered. The reactivation of latent pathogens and the acceleration of T-cell exhaustion for the second pathogen can explain several puzzling aspects of viral epidemics, such as COVID-19 and their unusual comorbidity mortality rates and post-infection symptoms.
T-Cell exhaustion can be referred sometimes as suppression indicating that certain types or classes of immune cells do not function properly. One of the hallmarks of this state is the "malfunction" of CD4+ and/or CD8+ T-Cells, which are among the most crucial components in the fight against infections, diseases, and cancers. This state can be induced not only by infections but also by long-term inflammation, which I often refer to as "long-term low-grade inflammation." Prolonged inflammatory conditions can over-stimulate the body, giving rise to some of the metabolic traps we discussed earlier, and can lead to the production of specific proteins that contribute to this state.
The T-cell inhibitory receptors include the programmed cell death protein 1 (PD-1), lymphocyte activation gene 3 protein (LAG-3), CD244, CD160, T-cell immunoglobulin domain and mucin domain-containing protein 3 (TIM-3), and cytotoxic T-lymphocyte-associated protein 4 (CTLA4).
T-cell exhaustion is reversible and/or can be avoided by blockades of specific co-stimulatory and inhibitory T-cell receptors, including a blockade of the interleukin-10 (IL-10) receptor, a blockade of the PD-1 receptor on CD8+ T-cells, and a blockade of CD4+ T-cell signaling by means of type I interferons (interferon α and β) Type I interferons α and β are essential cytokines in the early stages of viral infection for the activation and differentiation of CD8+ T-cells, but a long-duration release can induce CD4+ T-cell exhaustion
CD4+ T-cell exhaustion results in more CD8+ T-cell exhaustion, because of a reduction in interleukin-21 that is secreted by CD4+ T-cells And decreased functionality resulting from CD4+ T-cell exhaustion will reduce interferon-γ levels, essential for mobilizing intracellular defenses against acute and chronic pathogen infections.
I feel the need to directly cite this paragraph given what the first section states, which is in line with current (2023) evidence, T-Cell exhaustion is reversible by modulating (meaning using nutrients at a highly sufficient level or specific enzymes/proteins/drugs to influence) the receptors inducing the dysfunctional state, the “problem” is precisely targeting some of the receptors sometimes involved in the exhaustion process. The best method is just avoiding it and correcting the overactive immune response, especially if it is persistent, being present over a long period (months/years).
Human physiology is a perfect example of a highly complex but incredibly efficient system, with thousands of non-linear dynamics, and network effects, and as such the examples in the quote above cite these are not the only pathway towards suppression (dysfunction) or exhaustion of a important arm of your immune system, these effects have other pathways where many different proteins may interact with the ones cited above or other and create new cascade effects, a primary example is Galectin-9 (sister of 3) interacting with TIM3 and PD-1, and its high presence is correlated with poor cancer outcome. Galectin-9 “production” (expression) is promoted by Interferon beta and Gamma.
IFN-gamma is an old acquaintance of my readers, and also the protein that plays a significant role in the IDO (Kynurenine Pathway), is also directly responsible for this dysfunction/exhausted state. Using a very reductive form to frame it, T-Cell exhaustion could be described as a disproportionate state of anti-inflammation while the body still maintains inflammatory activity and damage, creating a “doom loop”. Another example of a “doom loop”.
T-cell exhaustion and Long COVID
A long duration of lymphocyte exhaustion for T-cells and NK-cells was also seen in COVID-19 survivors, even several weeks post-infection. Such multiple lymphocyte exhaustion could be a major causational factor for the COVID-19 post-infection symptoms collectively called Long COVID
In one case, a first pathogen infects a first subset of host cells and achieves T-cell exhaustion regarding the first pathogen, and a second pathogen infects a second subset of host cells and achieves T-cell exhaustion regarding the second pathogen. This situation would not facilitate reuse of the inhibitory ligands by the second pathogen, but the pre-existing inflammation and cytokine environment created in the T-cell exhaustion for the first pathogen could facilitate the second pathogen in more quickly achieving T-cell exhaustion. The delay time for the second pathogen to achieve T-cell exhaustion could be determined by these requirements: (1) the time needed for inhibitory ligands to be expressed by the host cell infected by the second pathogen, if not already expressed, and (2) the time (24 h–72 h) required for the T-cells for the second pathogen to express several inhibitory receptors
If the host is able to survive the accelerated T-cell exhaustion for the second pathogen and a faster-paced second pathogen infection from the weakened T-cell functionality, the first pathogen is a threat. If the first pathogen was a latent infection, T-cell exhaustion for the first pathogen can enable a reactivation of the latent first pathogen infection. Such a reactivation could potentially have severe consequences (such as encephalitis, hepatitis, or myocarditis, in the case of toxoplasmosis by T. gondii). Mother-to-fetus transmission of T. gondii and other protozoan parasites, or other maternally transmitted pathogens, or oral infections from food and water later reactivated by pediatric viral infections, could plausibly explain the recent world-wide pediatric cases of hepatitis seen in children lacking the viruses that normally cause hepatitis, if the latent pathogen reactivations were facilitated by recent infections of COVID-19
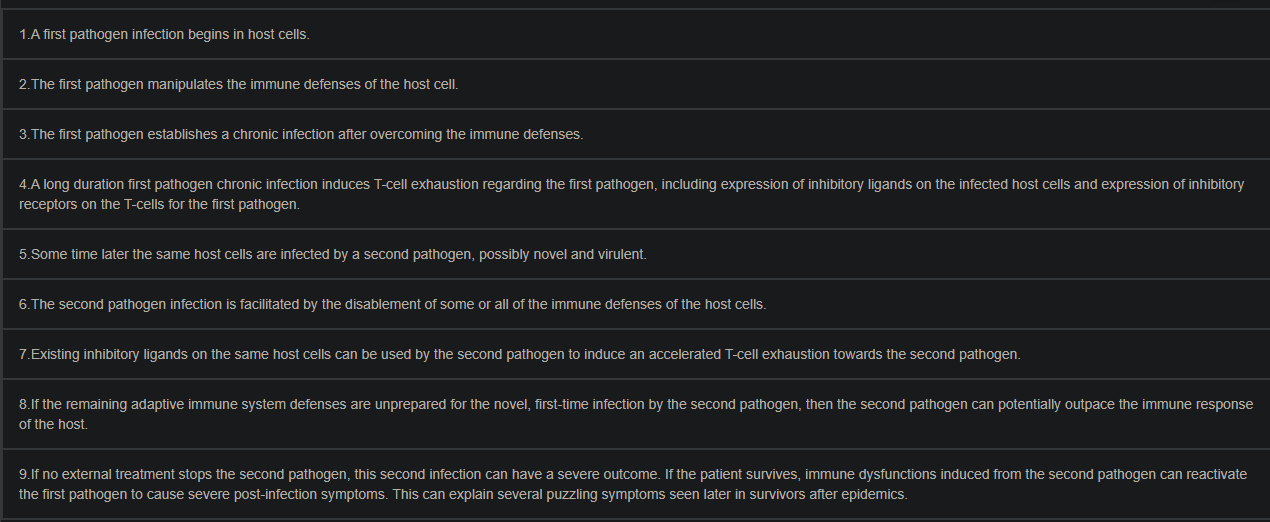
The author proposed model is clearly defined and is easy to understand for most people, and the entire citation above will be pertinent for years to come for a myriad of different reasons. And this is precisely how and why the long-term effects of both virus and spike protein are so complex because there are multiple, simultaneous feedback loops getting into gear which throws the immune system out of equilibrium.
The following scenario isn’t occurring population-wide, but it does happen in most severe and long-term symptomatic cases. By infecting your cells, SARS-CoV-2 creates a lot of ROS (highly unstable molecules, I like to call it cell rust) which depletes your antioxidant levels, by replicating given its heavy cysteine structure (it is made of a lot of small parts of cysteine) it depletes your cysteine levels, cysteine is the primary nutrient together with glycine used to make glutathione, the body most powerful antioxidant.
The following examples will be simplified, but the list of evidence is quite extensive on each of these.
This leads to mitochondrial stress, thus exacerbating T-cell exhaustion, hypoxia (a topic I covered a few times before) also plays a significant role. Uncontrolled oxidation leads to more inflammation thus initiating what we just described in this paper. Another aspect pertinent to all of this is the well-established role of sepsis and Covid, and sepsis has a well-established effect on both T-Cell exhaustion and immune paralysis. This state of immune paralysis increases the susceptibility to chronic and secondary infections, exacerbating CD8 exhaustion.
As back as May 2020, unbeknownst to authors, there were many hints of similarities between sepsis and Covid. A direct quote.
Such ineffective and detrimental expansions of innate/humoral responses, alongside T cell suppression, are reminiscent of classical features of sepsis, which is currently defined as a life-threatening organ dysfunction induced by dysregulated host response to infection, being characterized not only by systemic hyperinflammation (SIRS) with related endothelial and organ damage, but also by impairment of adaptive T cell immunity.
Here is a visual representation of T-Cell dysregulation in the critically ill.
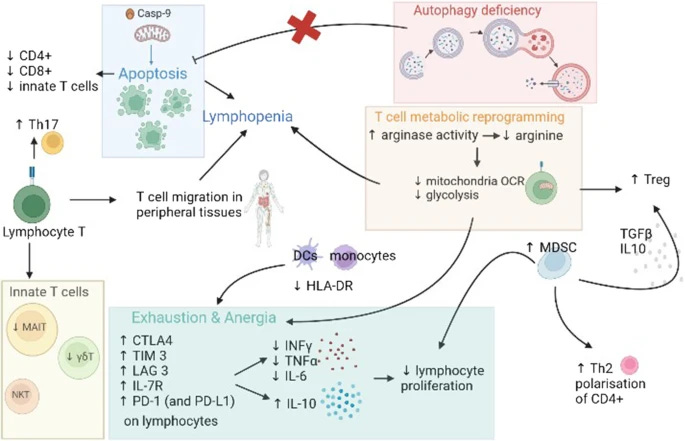
The complex states of T-Cell dysregulation affect at first your “naive” cells, cells that were not introduced to an antigen (you could call them baby T cells), so they can differentiate and enact their function. This inability of lymphocytes in general to recognize or respond to antigens is referred to as anergy. By affecting how CD8 cells behave and respond this induces a state of immune paralysis and exhaustion, and one of the consequences of this effect is the highly present reactivation of latent viruses in sepsis. Paradoxically enough, some of these effects such as the ones in the description of the image above can induce the body into both an allergic (Th2) state, an inflammatory (Th17) state, and as an attempt to control both induce immune suppression (T Reg).
One of the numerous reasons there is so much discussion and so numerous markers for different groups of Long Covid patients is that sometimes they find themselves in multiple contradictory (paradoxical is my preferred term) states.
The endotoxin (LPS) which binds to the Spike Protein is able to induce different alterations in lymphocytes function, and to understand how complex the situation in regards to virus, Spike, endotoxin is, the same protein TIM-3 that leads to exhausted cells, is also a novel negative mediator of endotoxic shock, meaning at first it is protecting the body, but the secondary and reactivated virus throw this into a paradoxical mess. One of the regions that can “glue” endotoxin to the virus is the Galectin-fold, which is a pretty close mimic of Galectin-3, it can also induce T cell exhaustion, which here is a problem.
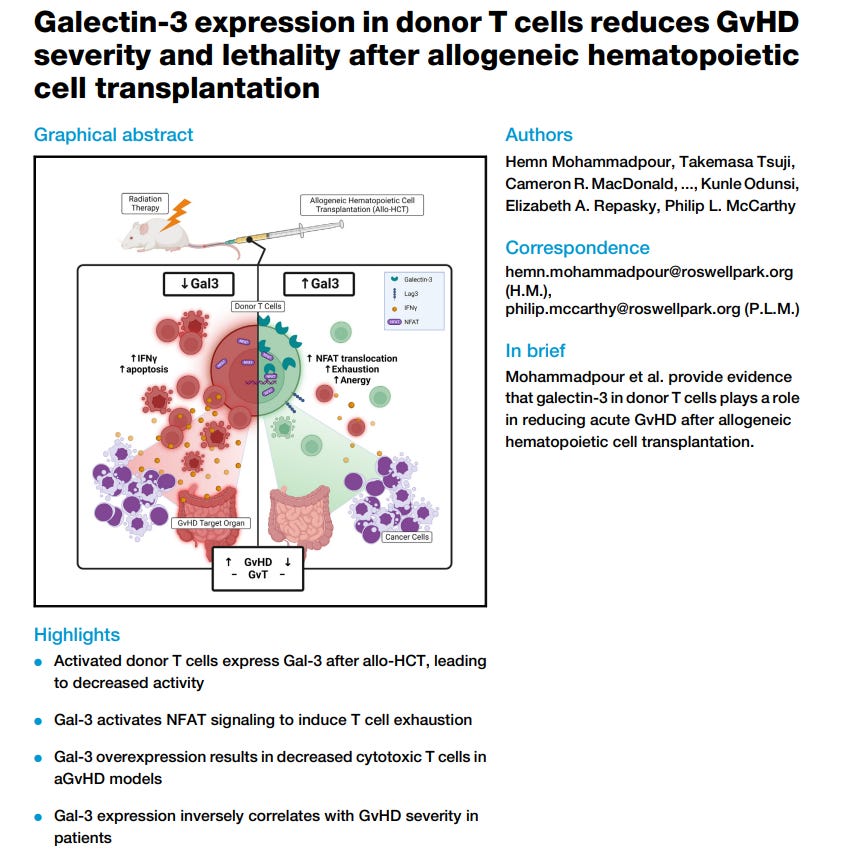
This a problem as it isn’t just a “twofer” but a multi-pronged effect, where each part on its own can already dysregulate the immune system, but together they act in synergy driving the effects further. And now we may be presented with another negative feedback loop when latent infections such as Herpes Simplex Virus 1 reactivation has higher levels of proteins and genes associated with T cell dysfunction and exhaustion. Same with Epstein Barr, which paradoxically even in this dysfunctional state (by how they measure it) can protect from the infection. The problem resides in the progression of said dysfunctional state leading to damage, inflammation, and therefore how some of these viruses are able to sometimes induce cancer.
One of the easiest and best solutions, presented by me and cited months ago remains the same, one is the cheapest method possible. Fasting. This will be the second time I cite this paper.

Fasting as key tone for COVID immunity a paper that cites adding BHB (Beta HydroxyButyrate) as an intervention method to recover immune function and immune cells. BHB is a byproduct of a ketogenic, or low-carbohydrate diet, and also a byproduct of fasting long enough.
Now you have a clearer picture of the complexity, and my somewhat insistence on covering a specific set of topics in a somewhat specific order, building towards this article. As a personal preference, I never edited older articles, as I prefer to “build” the work, also leaving a track record, as my readers can follow each step of the way. Therefore in line with latent viruses, sepsis, but outside of the current topic, I will add these.
A recent paper demonstrated myocarditis rates among the mRNA vaccinated are much higher than “experts” forecasted, a self-evident truth since 2021 anyhow, alas, I must bring this up, since the number of people with myocarditis being tested for what I am about to describe is close to 0.
Human herpesvirus 6-induced inflammatory cardiomyopathy in immunocompetent children
Cytomegalovirus (CMV) and acute myocarditis in an immunocompetent patient
Coxsackievirus myocarditis: interplay between virus and host in the pathogenesis of heart disease

I may write another T-Cell exhaustion article at some point in the near future. The following paragraph would be a better fit for the endotoxin paragraph, but it makes better sense as foreshadowing here.
Discussed by the authors of the seminal “The Spike Protein as an Endotoxin Delivery System” was the question of why or how the Spike Protein has this almost novel capacity to manipulate LPS, one of nature’s most powerful toxins to heighten its inflammatory-inducing capacity, and subvert the body responses. In the paper, they proposed perhaps the addition of the Furin Cleavage Site (FCS), the specific reason why SARS-CoV-2 is what it is and can infect humans with almost engineered perfection is the culprit, and perhaps an unintended consequence of the FCS. The FCS also resembles something else, one of nature’s most powerful proteins that are able to interact with our immune system.
A Superantigen, specifically the FCS, resembling SEB (Staphyloccocus Enteroxin B). I have never once mentioned superantigens in my Substack, but I will now, and they play a disproportional role in the long-term consequences of SARS-CoV-2. Especially a specific class of Superantigen that can interact with LPS and Herpesviruses.
I will be travelling from Friday to Sunday night, and it is the procedure of the business to have no electronics, so there will be complete comms blackout from me at some point from Friday until possibly Monday.
If i don't reply to your commentaries or questions in this article or older ones in the next few days, that is the reason. Same goes for e-mails.
I wish everyone a great weekend.
I asked you about SSRI’s and natto on Twitter earlier- it’s for my 21 yo niece who went behind my sister’s back and got two shots. She’s now having weird allergic reactions - can’t wear makeup anymore without eczema- like rash etc. Also just found out my 20 nephew has a newly enlarged heart- 3 x V. We tried so hard to reach all of our kids/ nieces/nephews, but they were under enormous pressure. Thankfully I kept mine safe, but lots more are at risk. Thank you for sticking your neck out to get us such important information and for personally responding. Safe travels. 🙏❤️