
The lasting consequences of SARS-CoV-2 fragments
Fragments part I... or II

In recent months, almost all my articles have been focused on a specific aspect of SARS-CoV-2, coming from a long-term perspective. The viral fragments and their consequences towards the body, especially microclots, what they are made of, how they are different, and all the different forms of damage they can induce.
But not only of clots, regardless of size, the damage and consequences are born from. Given the sheer complexity (no pun intended this time), not only from my work, but also from each paper, the fragment articles will be done in parts to avoid readers missing the significance of the subject being covered. The paper we are about to cover is a direct “follow-up” to the author’s previous work, which was both groundbreaking and remarkable. I will present a summary and explain the significance before going into it.
In the attempt to understand the lingering effects of SARS-CoV-2 and its consequential inflammation, the research team decided to use a machine learning approach to analyze the entire virus and its protein sequence and compare it to a specific class of peptides (very small proteins) we possess called Antimicrobial Peptides (AMP). They uncovered that certain sequences (small fragments after the virus and its proteins are broken down) in the virus mimic our own XMP, becoming xenoAMPs (xeno stands for foreign, not made in our bodies).
Many AMPs work by forming pores (holes) in the cell’s membrane, and by their very nature of fighting pathogens, are amylodogenic. The xenoAMPs produced by the breakdown of SARS-CoV-2 produce small clumps of protein that are both highly inflammatory and long-lasting because the body has a hard time breaking them down. Their persistence, a signal that lasts months, can cause systemic low-grade inflammation and thus can induce immune dysregulation.
While the infamous cytokine storm, when your body produces an absurd amount of inflammatory proteins as an immune response in a short time, remained the “poster child” of Covid research for years, another focus and poorly understood aspect persisted. Immune dysregulation. Both acutely (during the infection and shortly after) and long-term. How and why SARS-CoV-2 causes immune dysregulation, and to me especially, how it can cause immune cell death up to 10 months after an initial, mild infection, are central to understanding the overall “pandemic arc”.
SARS-CoV-2 peptide fragments selectively dysregulate specific immune cell populations via Gaussian curvature targeting
The authors cite Wuhan-era data on how many virions a Covid infection produces, billions to hundreds of billions of virions, virions being a fancy word for viral particles containing the viral RNA, its nucleocapsid protein, its envelope, and the infamous Spike Protein, thus an infection produces an abundant number of viral proteins. Theoretically, all these proteins are broken down by the immune system, be it as its normal cleaning process or for epitope generation, the little Lego blocks that initiate an immune response.
In their first test, they test how SARS-CoV-2 Spike Protein, both full-length and the S1 section, is processed by three different proteases (enzymes that break proteins down), because each one has a distinct specificity, they preferentially break down different sections of proteins. They use Lys-C, which mimics the broader family of trypsin-like enzymes, Elastase, which is derived from neutrophils, monocytes, and mast cells, and targets hydrophobic amino acids such as alanine, glycine, and valine. Lastly, Cathepsin G, which preferentially breaks down amino acids at their aromatic ones such as phenylalanine, tyrosine, and leucine.
All 3 proteases break down SARS-CoV-2 Spike Protein into short peptide fragments, but each protease produces distinct peptide fragments with different sequence coverage. Lys-C releases the least number of peptides, reaching only 25.8% of the original protein. Digestion by Elastase and Cathepsin G yields more peptide fragments with higher coverage of the original protein sequence, respectively being 42.2% and 43.8%.
But Lys-C generated peptide fragments have a stronger affinity to phosphatidylethanolamine (PE)-rich lipid membranes, which is a behavior seen often in AMPs. Fragments created by Elastase and Caghepsin G digestion lacked this strong affinity. To assess the potential of the many peptide fragments released during protease digestion (the breakdown of a big protein), they used a machine learning-based AMP classifier.
The classifier gives a score to measure the “AMP-ness” of each sequence, and results (E in the image above) show that all three proteases are capable of generating xenoAMPs when breaking down the Spike protein. For the trypsin-like protease Lys-C 8 of 26 fragments (30.8%) scored >0.4, which translates to 79.4% probabilityof the sequence functioning as an AMP. For Elastase, it was 3 of 74 (4%), and for Cathepsin G, 3 of 62 (4.8%). These are substantial numbers.
But not only of computer vibes, my funny way of saying computational modelling and analysis studies are made of. Since transmembrane pore formation (or forming roles in the cell membranes) is an established bactericidal mechanism of AMPs, they selected 3 peptide fragments as a byproduct of Lys-C digestion to determine if the xenoAMPs act similarly to natural AMPs. Each xenoAMP-fragment was incubated in a mixture that mimics cell membranes with high PE content very well. High PE promotes a negative curvature, the cell membranes bend inwards.
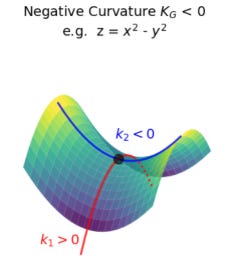
An explanation before the next part. A Negative Gaussian Curvature (NGC) is when a surface bends in opposite directions at one point, like this. Looks like a potato chip.
The results:
xenoAMP(538-558) (σ = 0.60): Induces a Pn3m cubic phase with lattice constant a = 18.2 nm (K = −0.020 nm⁻²).
xenoAMP(815-835) (σ = 1.19): Induces a Pn3m cubic phase with a = 19.7 nm (K = −0.017 nm⁻²).
xenoAMP(948-964) (σ = 0.34): Induces both Pn3m (a = 17.9 nm, K = −0.020 nm⁻²) and Im3m (a = 22.9 nm, K = −0.009 nm⁻²) cubic phases.
Bicontinuous cubic phases such as Pn3m and Im3m are lipid “shapes” where the bilayer forms a minimal surface with a saddle-shaped curvature at every point. In this topology, the change in the “cell membrane terrain” induces the formation of pores. The authors go further and quantify the negative Gaussian curvature and estimate the pore diameters to be in the range of 1.46 to 1.60 nm, comparable to canonical AMPs, and large enough to compromise membrane integrity and induce cell death. XenoAMPs not only affect the cell surface shape, but it is strong enough to start forming small holes.
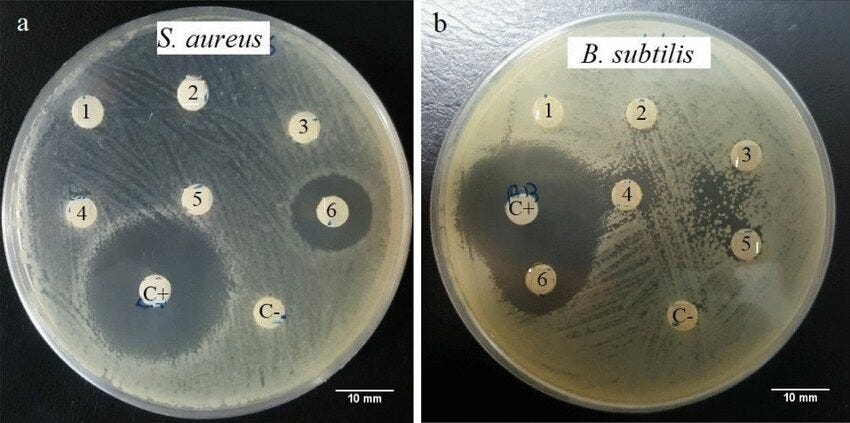
The next step is one of the most remarkable ones so far in the paper. By using Radial Diffusion Assay, where whatever you are testing will form circles as seen above if it is effective. They test and confirm that xenoAMPs possess genuine broad-spectrum antimicrobial activity against Acinetobacter baumannii, Enterococcus faecalis, Klebsiella pneumoniae, and Candida Albicans, with potency similar to our own AMPs.
Due to the chaotic nature of protein breakdown and subsequent diffusive (chaotic, random) transport, and also how inflammation can induce the body to produce its own “professional” AMPs, the authors wanted to examine whether a mixture of xenoAMPs and also a mixture of xenoAMPs and non-xenoAMPS, such as LL-37, could remodel the membrane.
First, they mix two different xenoAMPs, one high-scoring and xenoAMP fragment 445-562, and observe that they induce a membrane curvature larger than either can do alone. This means you don’t necessarily need high concentrations of specific fragments to generate pores. A mixture of xenoAMPs fragments will exert a similar effect similar to AMPs.
If XenoAMPs can synergize with each other, the next inquiry would be if pre-existing AMPs (as inflammatory conditions will create them) can do the same. In this step, they used LL-37. They found that viral xenoAMPs and LL-37 can induce pore formation via a unexpect mechanism they called “division of labor”. How it works in a simplified manner. LL-37 on its own is not capable of causing a shift in the cell membrane (to form pores), the same can’t be said for xenoAMPs. The more added, the stronger the effect.
But adding a normal amount of xenoAMP is enough to increase the concentration of LL-37, which then leads to a drastic increase in the pore formation. This effect is actually stronger than just adding more xenoAMPs by itself. In regards to SARS-CoV-2, the most likely source of LL-37 is neutrophils and their extracellular traps, because of how abundant they are in any Covid infection. The last step in this section is the most relatable to an actual infection.
Given that a natural infection will produce viral fragments covering a diverse range of sequences (each sequence is a fragment that covers a specific section, an example fragment covering 430-468, etc), they digest the intact spike protein into a mixture of peptide fragments by trypsin. Fragmented spike protein and intact spike protein are then mixed together. This experiment was done with a significantly lower amount of protein than the other experiments using xenoAMPs (1/8,000 ratio to be precise), thus the assortment of fragments generated during infection is outstandingly potent.
They did a parallel experiment with another trypsin-like protease called KLK5, significant because it is one derived from epithelial cells, and got similar results. These tests are often done with only one of these proteases and achieve the described results. There are multiple sources of proteases to create the viral fragments that end up acting as xenoAMPs.
So how do xenoAMPs know to target cell membranes with high local NGC (saddle, potato chip shape) ? Why are monoctyes, macrophages, and neutrophils spared while activated dendritic cells and T-Cells are targeted and end up dying ?
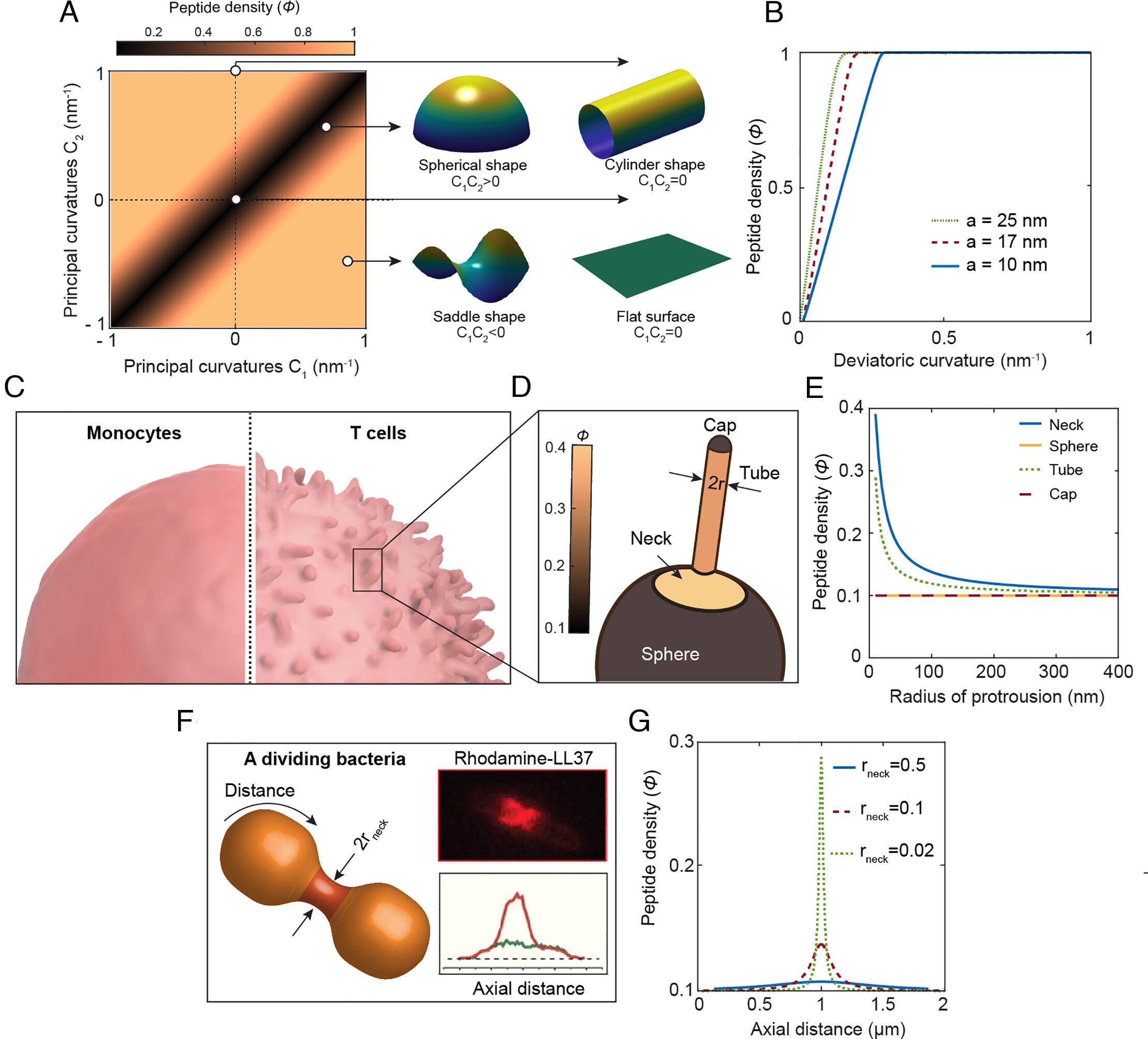
This section of the paper is both highly technical and complex (with a nice equation from the authors to explain their hypothesis and these effects, which can be expanded to so much else). I will oversimplify, as the analysis is dense at this point.
In this hypothesis, the author models the cell membrane as a thin, elastic sheet through which xenoAMPs can travel freely. A spherical or flat surface doesn’t have the necessary curvature (shape) for xenoAMPs to accumulate, let alone cause the membrane enough stress to form a pore, a saddle/potato chip shape does. But aren’t most cells spherical ?
Correct, so the distinction is shape. Our primary immune cells, such as monocytes, macrophages, and neutrophils, are mostly spherical and smooth-shaped, they have close to 0 of the necessary curvature for xenoAMPs to accumulate in these cells, so they accumulate far below the necessary amount to form pores.
On the other hand, activated dendritic cells and their processes have tentacle-like, spiky protrusions. And activated T-Cells have an abundant number of microvilli, tentacle-like projections of the cell membrane. Both of these are often modelled as a cylinder, and where the tentacle-like projection meets the membrane, called the neck, has a high negative Gaussian curvature, the more microvilli a cell possesses, the richer it is in NGC domains.
For these xenoAMPs, the tentacle-like projections and the high-curvature necks act as a trap, allowing even a lower amount of peptide to reach the cell saturation point, compromise its membrane, and form pores. And they verify their hypothesis. They apply this same curvature-based framework towards bacteria by using a human AMP.
The antimicrobial peptide LL-37 usually targets the septum of bacteria, here specifically E. Coli. The dividing bacteria form a constricting neck that is shaped like a funnel (catenoid-shaped), incidentally high on NGC. Per F in the image above, LL-37 strongly concentrates in the septum, at the funnel shape. This is the mechanism by which LL-37 arrests bacterial growth and subsequently compromises the cell membrane, forming pores, and inevitably killing it.
The last two sections of the paper provide more answers. SARS-CoV-2 Omicron Spike Protein has dozens (37) mutations, primarily concentrated at the NTD and the RBD, so the authors wanted to assess if Omicron’s fragments can induce a similar effect on cell membranes. Omicron’s viral fragments have a diminished capacity to induce NGC (the potato chip shape change) on the cell membrane. It still generates pores, but they are smaller and less stable, reducing the capability of Omicron xenoAMPs to induce cell death.
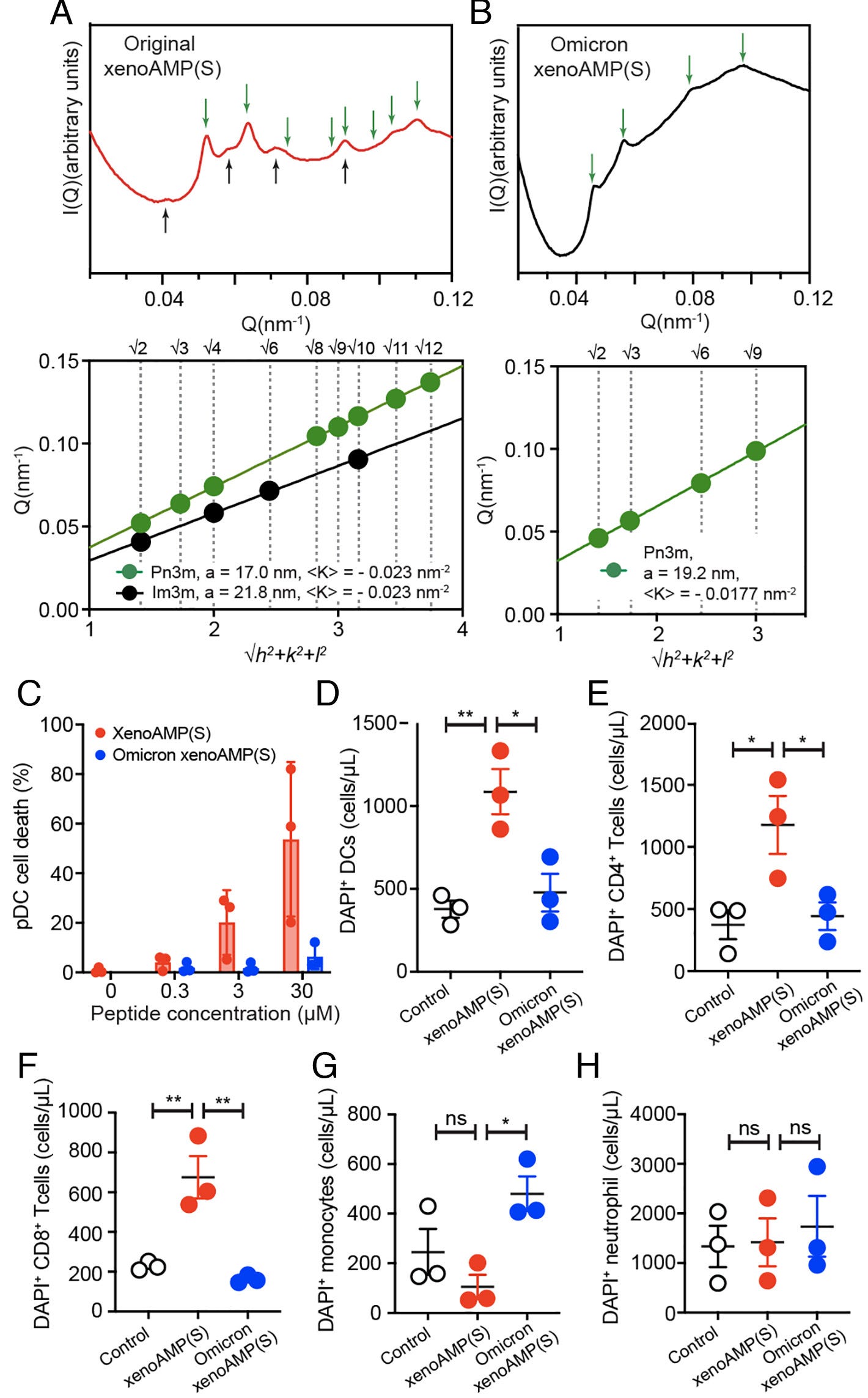
Lastly, they test whether SARS-CoV-2 xenoAMPs can permeate the membrane of immune cells and cause cell death. They isolated six immune cell types from the plasma of healthy donors. Original Wuhan strain xenoAMPs caused:
5.2 times (0.3 µM), 25.7 times (3 µM), and 68.3 times (30 µM) death of plasmacytoid dendritic cells, in comparison, Omicron xenoAMPs induced 8.1-fold cell death increase at a peptide concentration of 30 µM.
Dendritic cells 3.2 times dead cells
6.4 times for CD4+ T-Cells
5.4 times for CD8+ T-Cells
For these 3 types of immune cells, Omicron xenoAMP didn’t produce strong toxicity… with the exception of monocytes
The results from their tests on immune cells isolated from healthy donors provide clear evidence and replicate all the steps and findings in their paper. Immune cells with more drastic morphological projections, such as tentacle-like protusions from their cell surfaces, display more negative Gaussian curvature and are more sensitive to xenoAMP disruption and mediated cell death.
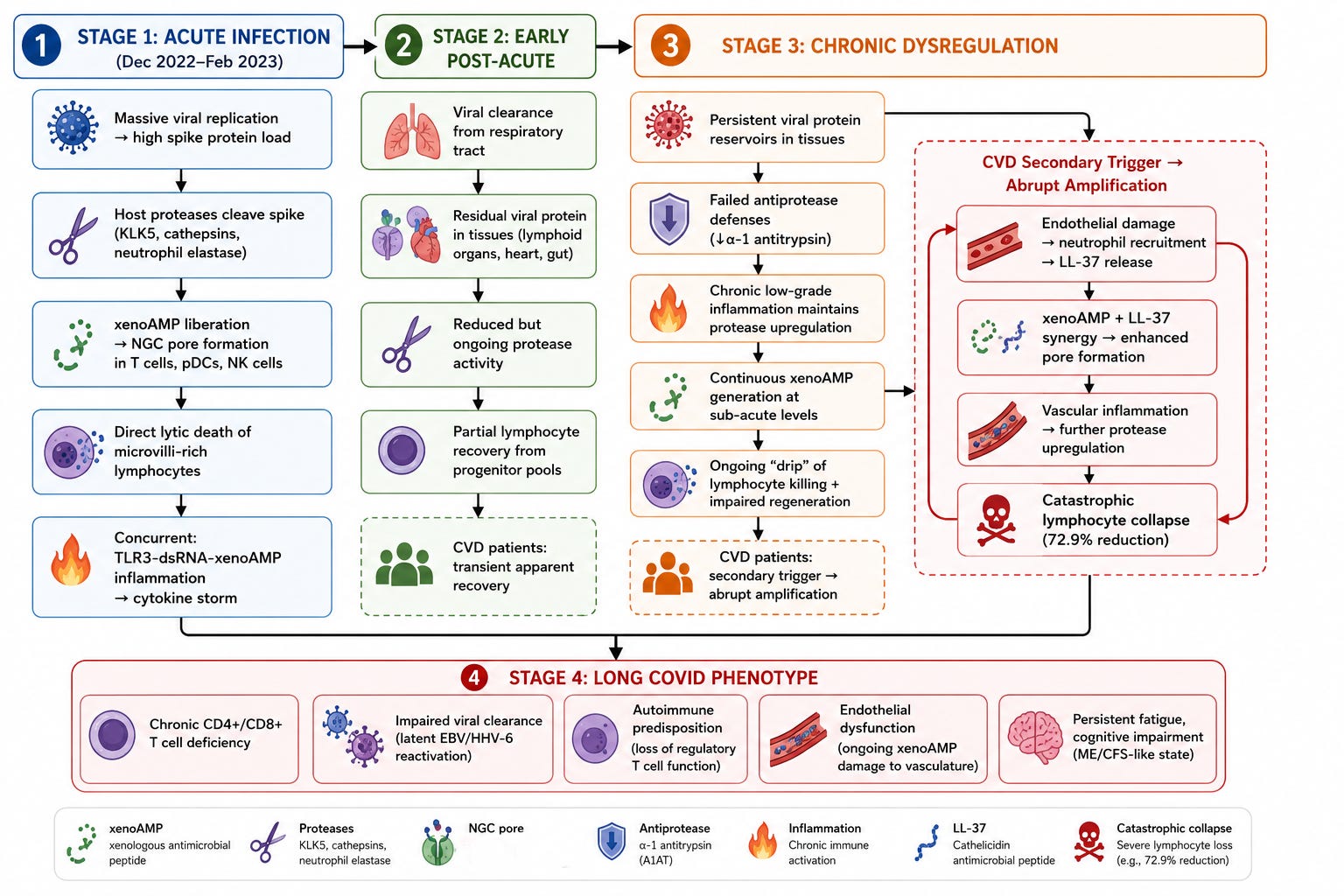
Their previous paper demonstrates clearly that xenoAMPs will interact with double-stranded RNA into ordered complexes that create a feedback loop of persistent inflammation for months, since these complexes are resistant to being broken down. This inflammation can lead to the activation of proteases (especially trypsin-like ones), which in turn leads to increased xenoAMP creation, which boosts inflammation and cell death.
There are a few studies demonstrating SARS-CoV-2 induces elevated protease expression, while, in parallel, a drastic decrease in antiprotease activity. This results in a state that can increase the production of viral fragments. This is a good map for understanding the long-term consequences of what was discussed here, with over a dozen of my articles providing deep analysis and data regarding the pathways in the image.
Going beyond Ultra Plus
The title above is an anime joke. Before integrating this remarkable paper into my own writing and observations. Omicron is likely toxic to monocytes by secondary mechanisms, while here they test xenoAMPs, we have ample evidence that the inflammatory and immunological conditions set off by even a mild infection lead to a significant shift in the immune system. No better way to describe than death by a thousand cuts.
To start, this paper finally provides clear evidence of what became one of the pillars of my primary hypothesis, called Paradoxically Acquired Immune Dysfunction. This demonstrates why there is a significant and oddly lasting drop in specific immune cell types regarding the SARS-CoV-2 mRNA vaccine, specifically in the CD8+ T cell population, and now provides a framework for why the drop lasts longer than it theoretically should. It also provides an answer to why exactly vaccinated people have such severely oxidative stress burden. Not only is the body attempting to dissolve the misshapen spike protein, but it is also entering a feedback loop created by its fragments.
We also now have evidence that the Spike Protein has not only one, but two sections that act as one of the world’s most powerful toxins, Staphylococcus Enterotoxin B (SEB). Both SEB, a superantigen capable of stimulating massive amounts of T-Cells quickly (it also, as a byproduct, increases the number of the tentacle-like microvilli), and the xenoAMPs reside in domains that are broken down by the proteolytic process.
During this process of superantigen-driven T-Cell activation, a lot of inflammation is released, which in turn causes endothelial damage, recruiting neutrophils to the site. Neutrophils then release their extracellular trap, which releases LL-37. This induces the increase of the proteases covered in the paper, creating more fragments. This mechanism can explain why susceptible people experience “odd events” weeks after an infection.
Both biological (via a secondary infection), mechanical (physical exertion, exercise), and emotional stress will induce the body to release viral fragments, which, during any of these events, will be broken down, creating an inflammation-driven immunological feedback loop. But the most impactful aspect of this paper to me was the fact that xenoAMPs have antimicrobial effects. Capable of killing both bacteria and fungi. Why is it so important ?
When xenoAMPs encounter bacteria, be it anywhere in the body, or bloodstream, they will compromise the bacterial membrane via the same pore-forming mechanism we observed in immune cells. This means that for gram-negative bacteria, it will release endotoxin, but also the contents in the interior, peptidoglycan fragments, lipoproteins, and bacterial DNA.
This will inevitably cause a temporary shift in the microbiome of any infected individual if they are healthy, but often via distinct mechanisms (direct viral interaction with the gut), it will compromise the gut barrier integrity, causing bacteria to translocate, creating a new flux of LPS when these bacteria come into contact with xenoAMPs.
The Spike Protein fragments themselves can carry LPS, but any cell that has its membrane compromised has enough space (because of the pore formed by the xenoAMP) to bypass normal LPS entry. This bypasses the canonical (well-studied) endotoxin pathway and uses the non-canonical inflammasome.
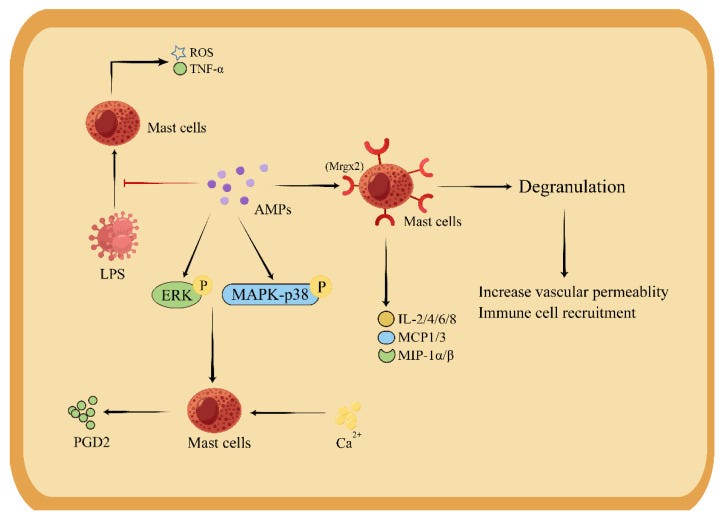
It is important to note how important the “division of labor” between xenoAMPs and LL-37 is. AMPs by themselves have profound effects on immune function. It has a dual function in macrophages, being able to produce the exact cytokines that have opposite roles (Th1, Th2). But more remarkably, it has direct effects on mast cells.
And while I chose to leave this for another opportunity, at the core of these mechanisms, becoming a sort of architectural central piece is HMGB1. The One Protein to Rule Them All. As there are other articles covering distinct fragments from the virus, I will have other opportunities to expand on this specific aspect.
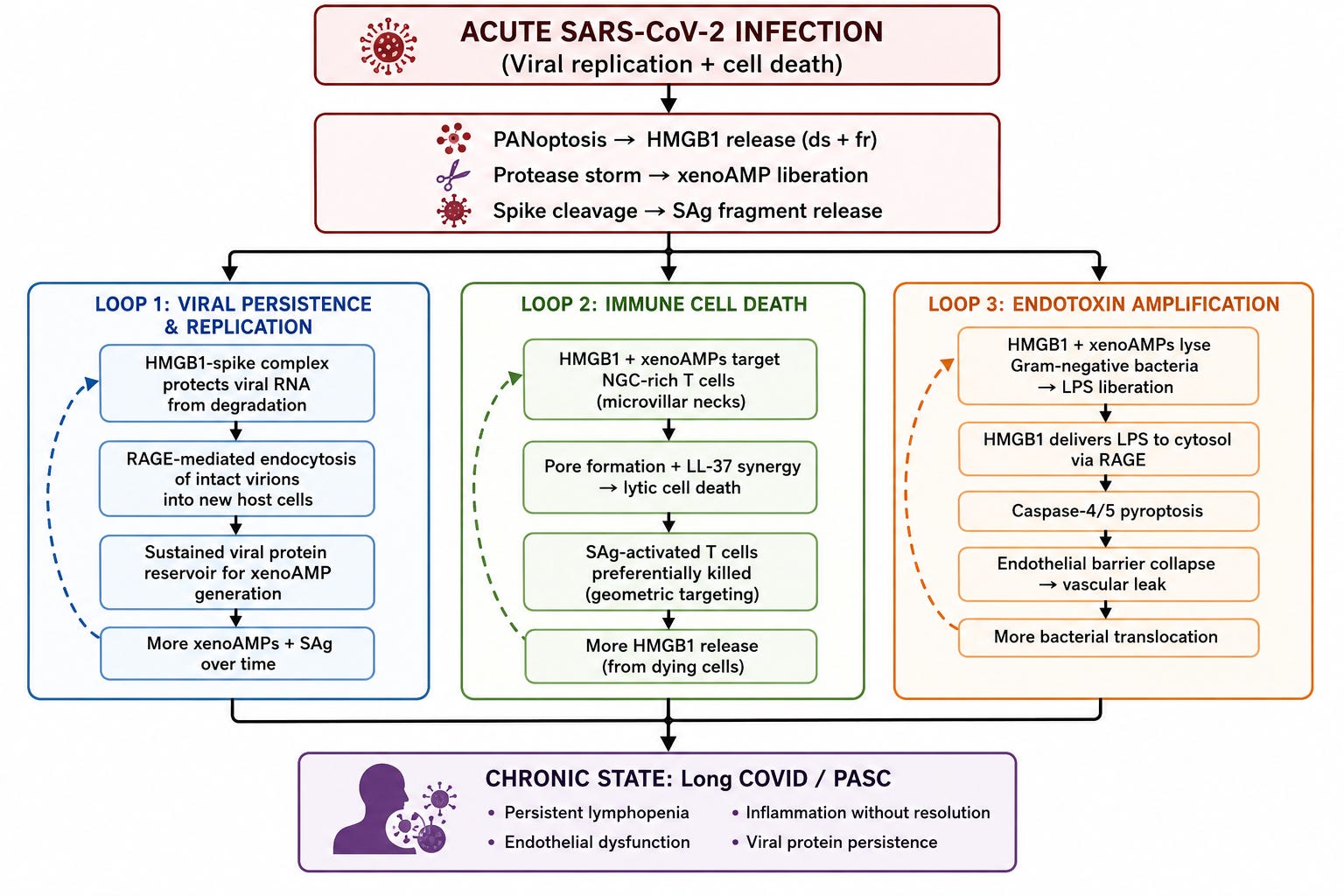
Below is a visual representation of all that has been discussed here.
Consider becoming a paid subscriber. As an alternative, you can buy me a coffee as a one-time thing

Any error will be fixed on Saturday or Sunday. There are always a few that seem to go through. I tweeted this on May 5.
"I am not one to believe in personal conspiracy, or think I have some importance in the world.
I have been without internet for days. Called the ISP technician. Usually it is an easy fix on the ISP side (sync stuff).
This time, someone literally cut my fiber optic cable. Only mine.
You can't achieve that without a work ladder and actively knowing which cable mine is.
For the first time in years, I actually think someone wants me to miss something.
What event or data point is the question."
And as a reply to someone.
"That is not a coincidence. The first words uttered by the technician were my same thoughts.
"There is no reason to cut fiber cable. There is nothing of value inside them." That is why fiber is widespread here, no resale value for metals."
To reiterate, I don't give myself any importance, I don't think I am important (my research, perhaps, may be helpful to people long-term, but that is). This is the first time active disruption has happened in my life.
Despite being unvaxxed, my blood is full of amyloid. I suspect the 5 covid infections I have had since Dec 28 2021 must be the reason. I appreciate your work tremendously. I had not seen either of these papers and this answers so many of my questions as to why I am plagued with the garbage in my system. 🙏 The cationic nature of these fragments also explain all the rbc rouleux formations in my blood smears. Nearly all rouleux, no normal single rbcs to be found. Despicable bastards.